

Physicochemical characteristics controlling the flammability of live Pinus banksiana needles in central Alberta, Canada
Rodrigo Campos-Ruiz A B * , Marc-André Parisien C and Mike D. Flannigan A B
A B * , Marc-André Parisien C and Mike D. Flannigan A B
A Department of Renewable Resources, University of Alberta, Edmonton, AB, T6G 2R3, Canada.
B Canadian Partnership for Wildland Fire Science, Department of Renewable Resources, University of Alberta, Edmonton, AB, T6G 2R3, Canada.
C Northern Forestry Centre, Natural Resources Canada, Edmonton, AB, T6H 3S5, Canada.
International Journal of Wildland Fire 31(9) 857-870 https://doi.org/10.1071/WF22008
Submitted: 29 January 2022 Accepted: 19 July 2022 Published: 25 August 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Background: Few studies have focused on the integral assessment of live fuel flammability in the boreal forest.
Aims: We aimed to examine the flammability of living needles of jack pine (Pinus banksiana) as characterised by their form, moisture and chemical content at different ages and times of the year.
Methods: With a calorimeter and open flame, we estimated needle ignitability, consumption rate, maximum speed and amount of energy released. We measured their form, moisture and chemical content.
Key results: Needle form has a major effect on ignitability, whereas chemical composition primarily influences the amount and rate of energy release. Needles <1 year old are the least flammable; they are rounder and voluminous, with higher moisture and nitrogen content. Needles ≥1 year old are more flammable; they are drier, more curved, thinner, longer, and contain more carbon and terpenes. Needles release more energy during the early growing season, when starch and lipids are at their peak concentrations.
Conclusions: Moisture content is not the major factor affecting the flammability of jack pine live needles; physicochemical changes specific to age and month of collection are the most influential factors.
Implications: Assessing the multi-faceted properties of live fuels flammability will help to comprehend stand- and landscape-scale fire behaviour.
Keywords: boreal forest, calorimetry, flammability, forest fires, jack pine, live fuels, pine needles, wildfires.
Introduction
Research on the flammability of fuels has been a priority for understanding forest fire behaviour for decades (Bond and van Wilgen 1996). Although tree crowns represent a significant source of fuel for high-intensity crown fires (the most prevalent fire type occurring in the Canadian boreal forest; de Groot et al. 2013), much of this research has focused on downed woody debris and litter. Conversely, the flammability of the living fraction of the boreal vegetation remains relatively understudied (Rivera et al. 2012; Finney et al. 2013).
The spatial and temporal context determine the assessment of forest flammability. At the particle scale (i.e. leaves, twigs, and small branches) and short periods (seconds), flammability encompasses four parameters: ignitability (ability to catch fire); combustibility (the rate of energy release); sustainability (fuel capacity to maintain flaming combustion, and the amount of energy released); and consumability (velocity of solid fuel conversion into gases) (Anderson 1970; Martin et al. 1994). At this scale, it is possible to quantify flammability in laboratory settings accurately, but not without limitations; flammability studies are very diverse in terms of techniques, equipment, fuel units and flammable parameters considered (Popović et al. 2021), which makes generalisations and comparisons challenging.
Foliar moisture persists as the primary perceived driver of forest flammability because of water’s strong physicochemical properties: high specific heat and high thermal conductivity. Consequently, water-rich fuels require larger amounts of energy during the preheating stage, so that water can evaporate and dissipate before they can burn. Additionally, water’s diffusivity as aerosol or steam reduces the immediate oxygen availability necessary for combustion (Byram 1959; Simms and Law 1967; Philpot 1970; Rothermel 1972; Pyne et al. 1996). Based on its known physicochemical properties and estimation simplicity, foliar moisture remains the cornerstone factor for forest fire modelling (Rothermel 1972; Van Wagner 1977; Alexander and Cruz 2012; McAllister et al. 2012). The high amount of moisture in living fuels led us to assume that they would burn similarly to dead fuels containing substantial moisture. In contrast to dead duels, living fuels can ignite before their moisture is expelled entirely (Pickett et al. 2010) and sustain and spread fire at foliar moisture contents >100% (Weise et al. 2005), whereas dead fuels hardly ignite or spread fire at moisture content >35% (Hawley 1926).
Dead fuels are functionally, physically, and chemically different from living fuels; therefore, their flammability differs. Dead fuels water dynamics follow relatively simple physical water absorption and evaporation principles, so they are more susceptible to the influence of weather. On the other hand, water uptake and losses by plants occur by evapotranspiration, a complex process involving their phenology, immediate environment, a multitude of interacting structures, chemical compounds, and other physical phenomena (Johnson 1966). Moreover, water in plant tissues acts as a solvent, providing colligative properties (Burgan 1979; Bloom et al. 1985; Raven 1998; Pyne 2007; Finney et al. 2013). Thus, understanding the physiological role and dynamics of water in plants helps understand their flammability.
Although multiple studies identify foliar moisture as the primary driver of flammability, their results also suggest interactions with organic compounds. Chemical components that function as structure (e.g. cellulose), defence (e.g. waxes, oils, and terpenes in the resin), and energy storage (e.g. fats and starch) are highly flammable (Carson and Mumford 1994), with the potential to enhance the flammability of the leaves. Despite results describing the potential effects of the chemical composition of leaves since the past century (Hubert 1932; Johnson 1966; Philpot and Mutch 1971), the study of flammability has, for decades, been predominantly focused on moisture content. More recently, researchers have turned to study how the chemical composition of leaves affect flammability, including fats (van Wilgen et al. 1990), terpenes and other volatile organic volatiles (Owens et al. 1998; Behm et al. 2004; de Lillis et al. 2009; Della Rocca et al. 2017; Romero et al. 2019; Ganteaume et al. 2021; Guerrero et al. 2021), sugars, starch, and assortments of these compounds (Jolly et al. 2012; McAllister et al. 2012; Page et al. 2012). Moreover, the shape and size of leaves regulate the heat transfer process and kinetics of combustion at the particle scale (Kanury 1994; McAllister et al. 2012; Naresh et al. 2018). Besides foliar moisture content, many morphological and chemical traits have been identified to affect flammability. However, an integral assessment of these characteristics related to the boreal tree’s phenology is lacking. This study aims to examine the flammability changes of living needles of jack pine (Pinus banksiana Lamb) ensuing from different developmental stages and needle ages during the growing season in central Alberta, Canada. We selected this species due to its abundance, proclivity to wildfires, and its role in the study of forest fire behaviour and ecology in Canada to develop the Canadian Forest Fire Rating System (Heinselman 1973; Little 1979; Van Wagner 1987; Gauthier et al. 1993; Farrar 1995). In particular, we aimed to:
Assess the relative contributions of living needle form, moisture and chemical content to their ignitability, combustibility, sustainability and consumability;
Identify the variables of needle form, moisture and chemical that drive the different aspects of flammability; and
Recognise the temporal patterns of needle flammability, form, moisture and chemical content during the growing season and according to their age.
Methods
To investigate the role of moisture content and jack pine living needles’ morphological and chemical characteristics on their flammability, we studied three different needle ages collected monthly during the growing season in Alberta, Canada. We employed a modified cone calorimeter with an open flame as the heat source and obtained the needles’ flammable characteristics: ignitability, combustibility, sustainability and consumability.
Sample collection
We collected needles from eight jack pine trees at the natural area of the University of Alberta Botanic Garden (Alberta, Canada, 53°24′07.1″N 113°45′14.6″W) between 1100 and 1400 hours each month, from June to September 2015. To reduce the environmental humidity influence on the needles, we sampled the trees on days with no rain over the previous 3 days. This site belongs to the Boreal Mixedwood ecological area, where jack pine is abundant in dry sandy soils (Beckingham and Archibald 1996). From June to September 2015, the average temperature and total precipitation normals in this area were greater for daily average temperature (14.8°C) and lower for total precipitation (221 mm) than the climate normals (1981–2010; 14.0°C and 303 mm; Supplementary Table S1, Government of Canada 2019). This period overlaps with the highest wildfire activity in Alberta, also referred to as ‘fire season’ (1 March–31 October), which exhibits a peak of number of fires during the summer (specially June and July; Campos-Ruiz et al. 2018).
We prepared the samples for flammability and biometrical analysis (form, and chemical) in the laboratory. For each monthly sample, we clipped three twigs from the middle section of the crown in the south-facing side of the same trees. We stored them in sealed plastic bags and transported them inside a cooler with icepacks to the laboratory, where we kept them in a fridge at 4°C. The needles were manually detached from the twigs and, based on their distance from the distal point of the twig, separated them according to their age: 0 years old or new (newly emerged during the current year), 1 year old, or 2 years old. We obtained 96 stacks of pine needles (samples) from the combination of three needle ages (new, 1 year old, 2 years old), 4 months (June–September), and eight tree replicates. We divided each sample into three subsamples destined for flammability, chemical and morphological analysis.
Flammability tests
We subjected the first set of subsamples (~4 g or 50 needles each) to an oxygen consumption calorimetry experiment employing a modified mass loss cone calorimeter (Mass Loss Calorimeter ISO 13927, Fire Testing Technology, East Grinstead, West Sussex, UK) at the Protective Clothing and Equipment Research Facility of the University of Alberta, Edmonton. The cone calorimeter measures heat release (EHC), heat release rate (HRR), and gravimetric changes of a sample exposed to a constant heat flux (Babrauskas 1984). The heat source is usually a radiant heater with the shape of a cone positioned over the sample. We opted for a more realistic setup where we exposed the samples to the radiation and convection of a flame instead of a radiator. We employed an open methane burner (10 × 10 cm) set at a constant flow of 9 L per min (Melnik et al. 2022; Supplementary Fig. S1). We estimated the time to ignition (IGT) as the time elapsed between the burner start and the observation of a flaming reaction of the sample. This time was later confirmed with the heat release rate output from the sample (Supplementary Fig. S2). We subtracted the contribution of the flame from the output to obtain the differential heat release rate (i.e. the difference between measurements with and without a sample, hereafter HRR) and the effective heat of combustion based on the total heat release (EHC; Table 1). We placed each subsample uniformly on a metallic mesh holder (10 × 10 cm) at 5 cm over the burner and exposed it to the flame for at least 80 s when the whole sample was consumed entirely. We interpreted the outputs in terms of flammability components: time to ignition (ignitability); peak heat release rate (combustibility); effective heat of combustion (sustainability); and the average rate of mass loss (consumability; Table 1).

|
Moisture content
We calculated the foliar moisture content (FMC, dry-based %) from the form subsamples by weighing the needles before and after drying them in a convection oven at 75°C for 48 h (Table 2).

|
Form analysis
We measured form characteristics from the second set of subsamples by scanning 20 needles per treatment combination and replicate (age × month × tree) and processed the images with the WINSEEDLE software (Reagent Instruments Incorporated, Quebec, QC, Canada). We obtained the sample averages (per needle) curvature and form coefficient and calculated the surface-area-to-volume ratio by assuming the volume of a hemi-ellipsoid and adjusting the projected area to a hemi-surface leaf area (Table 2; Bond-Lamberty et al. 2003).
Chemical analysis
We kept the third set of subsamples frozen at −25°C and sent in a cooler with dry ice to the Chemistry Services Laboratory (Victoria, BC, Canada) of Natural Resources Canada for chemical analysis. The analysis included the most abundant, functionally relevant and high energy components of needles: total nitrogen, carbon, non-structural carbon (NSC’s: starch and soluble sugars), lipids, and terpenes (Table 2. Detailed methods of extraction and measure in Supplementary Table S2).
Statistical analysis
The experiment was based on a repeated-measures factorial design to test the effects of age (three levels: 0, 1 and 2 years old) and month (June, July, August, and September) on their flammability, foliar moisture content, form, and chemical makeup of the needles. First, to explore and visualise the similarity between samples and possible relationships between flammability and biometric variables, we used a non-metric multidimensional scaling (NMDS) procedure. Second, to confirm the significance of the sample groupings observed with the NMDS, we used its scores to test for differences between ages and months of sampling with a permutational Multivariate Analysis of Variance (MANOVA) for repeated measures. Third, we calculated the relative contributions of moisture content, form and chemical content on flammability through variance partitioning, based on the NMDS output. Fourth, we ran correlations for repeated measures to confirm associations between flammability and biometric variables. Finally, we tested the effect of needle age and month of collection on each flammability and physicochemical variable, employing a repeated-measures analysis of variance (rmANOVA) and pairwise comparisons.
We calculated the distances between observations with Bray–Curtis dissimilarities (‘distance’ function, ecodist package in R; Goslee and Urban 2007; R Core Team 2008), ordinated those observations via the ‘nmds’ function (vegan package in R; R Core Team 2008; Oksanen et al. 2020), and grouped them by age with 95% confidence limit ellipses. To test whether age and month were significant, we employed the scores derived from the NMDS (two-dimensional) and used a permutational MANOVA for repeated measures (‘MANOVA’ function in the MANOVA.RM package for R; Friedrich et al. 2018). The power to detect significant differences among treatments by MANOVA was 0.72 (α = 0.05, n =96) (G*Power software; Faul et al. 2007). This value, which ranges from zero to one, reflects the capability to find differences between treatments based on alpha, the magnitude of expected change (medium in this case), the number of samples, the number of treatments, and the number of repeated measurements.
Next, we performed a variation partitioning procedure to determine the relative and combined explanatory power of moisture content and grouped morphological and chemical characteristics on each flammability component. We employed the ‘varpart’ function from the vegan R package using the grouped variables in Table 2. Finally, we used distance-based redundancy analysis (dbRDA) to assess the significance of the unique variation explained by each of the three groups.
We used repeated-measures correlation tests (RMCOR) and NMDS to find potential relationships between flammability variables with chemical and form features. RMCORR calculates correlation coefficients (rrm) in repeated measures (within-subjects) designs without averaging the repeated measures that may obscure meaningful relationships. The resulting coefficient delivers the same interpretation as a Pearson correlation coefficient, with the advantage of having higher degrees of freedom and power than averaged data analysed with traditional correlation techniques. Additionally, in RMCOR, the effect size is calculated with bootstrapping to estimate the parameters’ accuracy; thus, non-normally distributed data is supported. We calculated the correlations between variables using the ‘rmcorr’ function in R (rmcorr package; Bakdash and Marusich 2017).
We tested the effect of needle age and month of collection on flammability, morphological, and chemical variables with rmANOVA. We checked the normality data and residuals for each variable. If any of the assumptions for this test were violated, we transformed the data (log, root squared, or logit transform) before the analysis. We performed the rmANOVA (specifying for repeated measures on the subjects), Tukey multiple comparisons test, and grouping. These tests were performed with the functions ‘aov’, ‘lsmeans’, and ‘cdl’ in base R, lsmeans, and multcomp packages; (Hothorn 2008; Lenth 2016). We adjusted P-values through false discovery rate methods to control type I errors.
Results
Flammability
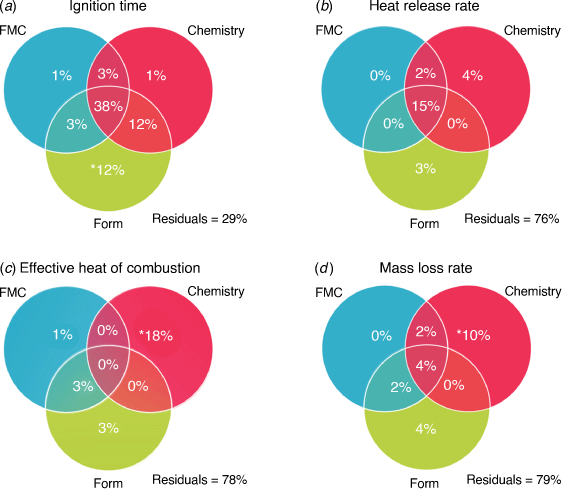
Younger needles, especially in the earlier growing season, were significantly different from older ones based on their flammability, FMC (foliar moisture content) and biometric variables, which results in distinct grouping in the NMDS (Fig. 1). Biometrical variables (form and chemical composition) explained most of the variance of each flammability component, but FMC explained the least (Fig. 2). The older needles (1 and 2 years old) had higher HRR (heat release rates) and MLR (mass loss rates) and lower IGT (ignition times) than new (0 years old) needles (Figs 1, 3a, b, d). We observed significantly higher EHC (effective heat of combustion) during the first half than the second half of the growing season, regardless of age (Fig. 3c). We found multiple significant correlations between IGT, HRR, EHC and MLR with the needles’ physical and chemical characteristics, such as foliar moisture content, form coefficient, curvature, surface-to-volume ratio, Nitrogen, Carbon and terpenes. Only EHC showed correlations with lipids and starch (Table 3). In general, age and month of collection had a significant effect on live needle flammability, their moisture content, form and chemical composition (Supplementary Tables S3–S5).

|
|

|

|
The resulting NMDS showed that new needles formed a distinct group almost independently from the rest of the data during June and July, but overlapped in August and September with the oldest age groups. At least one group resulting from age and month of collection in the NMDS (Fig. 1) was statistically different from the rest (rmMANOVA, P = 0.001, month P = 0.001, number of permutations = 999). The first axis reflects significant age differences, with variations in IGT, HRR, and MLR associated with FMC, FCO, CRV, AVR, N, C, and TE content. In contrast, the second axis reflects the month of collection and variations in EHC associated with ST and LI. All variables, except for SS, were significantly correlated to the ordination axes (Supplementary Table S6). The NMDS had a stress value of 0.05, which is an excellent representation of the data in two dimensions (Clarke 1993; Fig. 1, Supplementary Table S6).
Foliar moisture content, form, and chemical content
The contribution of foliar moisture content to flammability variance was minor compared with form and chemical composition. According to the variation partitioning, FMC only accounts for 1% or less of each component of flammability’s unique variance, whereas form and chemistry explain larger amounts (Fig. 2). Needle form, chemistry, and FMC explained 71% of IGT variance together. Although FMC, form and chemistry explained 45% (1 + 3+ 3 + 38), 65% (12 + 3 + 38 + 12) and 54% (1 + 3+ 12 + 38) of IGT variance respectively, a large percentage (38%) was explained simultaneously by the three sets of variables (shared or redundant). FMC and chemistry only explain 1%, whereas form explains 12% uniquely (Fig. 2a).
Chemical composition explained most of the HRR variance, shared and unique. Chemistry, form, and FMC explained 21, 18 and 17% of HRR variance respectively, but uniquely they each explained <4%. In total all variable sets explained 24% of HRR variance, from which 15% was shared (Fig. 2b). Chemistry also explained most of EHC and MLR variance (shared: 18 and 16%, uniquely 18 and 10%). Form and FMC explained the least variance for EHC and MLR (shared <8%, unique <4%; Fig. 2c, d). It is important to note that EHC variance explained by chemistry did not overlap (i.e. it is not redundant). Residuals for HRR, EHC and MLR ranged from 76 to 79% suggesting that other variables not included in our study may explain more of their variance.
Flammability components were correlated with most of the needles’ tested morphological and chemical characteristics. IGT had a strong negative correlation with AVR ratio (rrm = −0.42, d.f. = 87, P < 0.001) and positive with FMC, FCO, N and LI content (rrm ≤ 0.25; Table 3). Then, the more ignitable needles (i.e. low IGT) had higher AVR, were narrower, longer, drier, and had less N and LI. Peak HRR exhibited negative correlations with MC, FCO, N, but positive C, TE, and CRV (all with rrm ≥ 0.22; Table 3). EHC was positively correlated to starch (rrm = 0.33, d.f. = 87, P < 0.01) and lipid content (rrm = 0.31, d.f. = 87, P < 0.01). MLR had positive correlations with CRV, C, and TE content (Table 3), which means that needles consumed faster by fire were curvier and had more carbon and terpenes.
Needle age and month of collection
Old needles ignited significantly faster than new ones (rmANOVA, F = 45.69, d.f. = 2, P < 0.001) in the first 2 months of collection (rmANOVA, F = 36.24, d.f. = 3, P < 0.001), but especially in June (rmANOVA, F = 28.21, d.f. = 6, P < 0.001; Supplementary Table S3). It took up to 49.60 s (x̄, s.e. = 3.69) for the 0-year-old needles to ignite in June, whereas it occurred in less than 22 s for the 1- and 2-year-old needles (Fig. 3a). Differences in ignitability due to age disappeared in August when all needles ignited under 16 s.
The highest HRR was found in 1- and 2-year-old needles (rmANOVA, F = 16.19, d.f. = 2, P < 0.001), especially 2-year-old needles in July (x̄ = 130 kW m−2, s.e. = 10.41), and the lowest in 0-year-old needles in June (x̄ = 63.10 kW m−2, s.e. = 3.10). HRR was the lowest for all needle ages in June but reached their maximum value at different months: 1-and 2-year-old needles in July, and 0-year-old needles in August. Thus, month (rmANOVA, F = 11.23, d.f. = 3, P < 0.001) and interaction (rmANOVA, F = 2.80, d.f. = 6, P < 0.05) also had a significant effect on HRR; Fig. 3b, Supplementary Table S3).
The EHC was similar between needle ages (0, 1, and 2 years old) but varied depending on the month of collection (rmANOVA, F = 13.26, d.f. = 3, P < 0.05; Fig. 3c, Supplementary Table S3). During June and July, needles released almost three times more energy (x̄ = 11.30 kJ g−1, s.e. = 2.00) than in August and September (x̄ = 4.40 kJ g−1, s.e. = 1.05).
The amount of FMC was higher in 0-year-old than 1= and 2-year-old needles, mainly during the first 2 months of collection (Fig. 4a). New needles exhibited a maximum FMC of 295% (x̄, e.e. = 35.7) in June and dropped to 129% in September (x̄, e.e. = 4.51), a level similar to older needles (~100%). FMC of needles, then, was significantly affected by age (F = 94.47, d.f. = 2, P < 0.001), month of collection (F = 12.13, d.f. = 3, P < 0.001), and interaction (F = 13.99, d.f. = 6, P < 0.001).

|
Form and chemical characteristics
Age and month significantly affected the needles’ form (Fig. 4, Supplementary Table S4). New needles (0 years old in June) showed low curvature (x̄ = 0.30, s.e. = 0.002), high form coefficients (x̄ = 0.26, s.e. = 0.01) and low area-to-volume ratio (x̄ = 6.36, s.e. = 0.33), whereas the opposite was observed in older ones (Fig. 4b–d). Chemical content varied according to age and month of collection (Fig. 5, Supplementary Table S5). Needles’ growth during the growing season and years led to increases in carbon and terpene content but a reduction of lipids (Fig. 5a, e, f). We identified 50 different terpenes (36 monoterpenes, 12 sesquiterpenes and two diterpenes). The most abundant were α-pinene, β-phellandrene, borneol acetate, β-pinene, camphene, β-myrcene, 3-carene, linalool, germacrene D-4-ol, myrtenyl acetate, (+)-R-limonene and phytol.

|
Starch content in older needles decreased over the months, eventually matching new leaves (from 7 to ~0.21%; Fig. 5c). Lipids fell consistently for needles of all ages as the growing season advanced (Fig. 5e), soluble sugars had a pattern of highs and lows (Fig. 5d), and nitrogen decreased drastically in 0-year-old needles after June, matching the other needle ages. At the end of the growing season (September), the needles’ chemical characteristics were similar regardless of age.
Discussion
Ignitability, combustibility, sustainability and consumability reach their maximum at different times of the year and vary with needle age, matching both natural developmental and seasonal stages of jack pine needles. Needles ignite and release energy faster at maturity, which is also when they are no longer significantly different physically or chemically. However, the total energy released responds to the shift of needle chemical composition between the early and late growing seasons regardless of their developmental stage. During the early season (i.e. June and July), the energy is three times the amount released later (e.g. August and September), a difference similar in magnitude between living and dry, dead needles of P. halepensis (~10 kJ g−1; Jervis and Rein 2016). There is an intrinsic potential for more intense crown fires in the early fire season, which is also the period with the region’s highest number of forest fires. However, weather and topography also modulate fire intensity at larger spatial scales. Nevertheless, although more research is needed to study the interactions between live foliage flammability and larger-scale factors, our findings highlight the importance of flammability studies of living foliage in a biological context.
The form and chemical content of jack pine needles had a more substantial effect on flammability than foliar moisture content. We observed the lowest ignitability and combustibility at higher FMC, consistent with similar experiments testing conifers and broadleaved species (Etlinger and Beall 2004; Weise et al. 2005; Jervis and Rein 2016). However, despite their significant correlations with FMC, two observations showed that FMC was not the only factor driving flammability. First, needle ignition occurred before their desiccation (also described by Pickett et al. 2010), contradicting the traditional combustion progression (Byram 1959). Second, the combustibility does not precisely track the monthly patterns of needle moisture content (cf. Philpot and Mutch 1971). In fact, FMC in living conifer needles could contribute to the sudden release of droplets (micro-explosions) mixed with volatile compounds that can heat and ignite adjacent needles (Darwish Ahmad et al. 2021; Fazeli et al. 2022). Previous studies also acknowledged that leaves’ form and chemical content also affect flammability (Philpot and Mutch 1971; Weise et al. 2005; Jolly et al. 2012, 2016; McAllister et al. 2012; Jervis and Rein 2016). However, our analysis also allowed us to recognise the extent of influence of each set of variables on flammability and indicated a more minor role of FMC than we were expecting. These results suggest that under the conditions of a high-intensity crown fire, the effect of moisture content may be considered negligible on ignitability and combustibility (Weise et al. 2005; Fletcher et al. 2007; Alexander and Cruz 2013), or limited to ignitability (Ganteaume 2018) and the peak rate of energy release. This low contribution of FMC to flammable characteristics may explain the lack (or undetectable role) of this variable in crown fire behaviour models (Van Wagner 1998; Cruz et al. 2005; Rossa and Fernandes 2018). Given the preponderance of high-intensity crown fires in the Canadian boreal forest, performing more experiments on a broader scale and a larger number of species will help us determine the extent and type of influence FMC exerts on the flammability of living foliage of trees.
Moisture content may indirectly influence ignitability by modifying jack pine needles form. Needles with high moisture content are more voluminous and rounder, which increases their thermal capacity and delays ignition. This observation is similar to that of Jolly et al. (2016), who also noted delayed ignitability at higher needle mass and density, which in turn was associated with higher FMC and starch contents in red and jack pine. Such associations might also explain the high redundancy of variance explained by FMC and form. The tight association between form and ignitability has allowed researchers to rate different species by their fire hazard, and to develop fire spread and severity prediction models based solely on leaf form traits (Montgomery and Cheo 1971; Papió and Trabaud 1990; Weise et al. 2005; Murray et al. 2013, 2020; Shen and Fletcher 2015).
The chemical composition of jack pine needles is the main factor determining sustainability and consumability, and has a minor role in combustibility and ignitability. High needle carbon content boosts ignitability, combustibility and consumability. Carbon increment in new needles reflects the rise of lignin and cellulose, which form structural tissues (sensu lato structural carbons; Kozlowski et al. 1991). Although they reduce flaming times and energy release in lodgepole pine dry needles (Page et al. 2012), lignin is a source of high energy in the combustion of wood (White 1987). More importantly, non-structural carbon like starch and lipids are strongly associated with sustainability. These high-energy compounds observed in many conifers, are more abundant during shoot expansion in the early growing season, (Little 1970; Fischer and Höll 1991; Mandre et al. 2002; Hoch et al. 2003; Schoonmaker 2013) when their high concentrations may increase needles’ flammable characteristics (Philpot and Mutch 1971; Page et al. 2012; Jolly et al. 2016). This implies that jack pine needles have the potential to burn more intensely during the first half of the fire season.
Terpenes tend to get immobilised in the needle tissues over time in the resin canals (Langenheim 1994), increasing the flammability of older leaves. In our study, they enhanced the consumability, combustibility and ignitability of living jack pine needles due to their inherent high heating value, low ignition temperature (flashpoint), and low ignitable concentration (lower flammability limit; Carson and Mumford 1994). In several species, a significant relationship exists between some terpenes and ignitability (de Lillis et al. 2009; Pausas et al. 2016; Romero et al. 2019; Della Rocca et al. 2020; Guerrero et al. 2021), combustibility (Dewhirst et al. 2020), and consumability of foliage (Owens et al. 1998). Moreover, terpene content can explain high percentages of variance with respect to flammability (19–24%; Della Roca et al. 2017). However, their role depends on the flammability parameter studied, their type (number of isoprene units and associated functional groups in their structure), season and developmental stage (Della Roca et al. 2017; Dewhirst et al. 2020; Ganteaume et al. 2021).
In the present study, the most abundant terpenes in jack pine needles are monoterpenes, which also play a significant role in the flammability of living foliage of Globularia alypum and Rosmarinus officinalis (Alessio et al. 2008; Pausas et al. 2016), 13 species of pines (including P. banksiana; Dewhirst et al. 2020), and several Mediterranean conifer species (Della Roca et al. 2017; Ganteaume et al. 2021). Needles emit terpenes and other volatile organic compounds when exposed to heat (Greenberg et al. 2006), which may have accelerated and caused ignition prior to needle desiccation in our samples (Darwish Ahmad et al. 2021; Fazeli et al. 2022). We identified terpenes in our samples that are considered highly flammable (e.g. α-pinene; Raman et al. 2016), and others that are recognised as supressors of flammable properties (e.g. β-pinene and α-humulene; Ganteaume et al. 2021) in other studies. Nevertheless, the suppressing or enhancing role of individual terpenes on flammability was beyond the scope of the present work.
Both N and FMC are abundant in jack pine needles during shoot expansion due to the high physiological activity involving multiple proteins, enzymes, amino acids, and nucleic acids (Kramer and Kozlowski 1979; Vose and Ryan 2002). At this stage, needles are less ignitable and combustible than at later stages, but distinguishing the role between N or FMC cannot be asserted entirely. On the other hand, when only considering older needles, higher N concentrations might reflect different N fractions such as volatile flavonoids and phenolic precursors of lignin (Zakzeski et al. 2010), with highly flammable properties such as piperidines (Patnaik 2007; Virjamo and Julkunen-Tiitto 2016). Further research is necessary to assert the role of more specific compounds in the flammability of jack pine needles.
Although small-scale controlled experiments have been considered limited in their ability to explain field fire behaviour (Fernandes and Cruz 2012), the results obtained from them are valuable to cement our understanding of underlying factors influencing combustion. This information can potentially be used to improve mechanistic-based models of fire behaviour involving crown fire initiation, rate of spread, and energy release. These experiments have facilitated the rating of vegetation into flammable categories (Papió and Trabaud 1990; Dimitrakopoulos and Papaioannou 2001; Weise et al. 2005), the management of the wildland–urban interface (White and Zipperer 2010; Simeoni et al. 2012; Ganteaume 2018; Krix et al. 2019; Murray et al. 2020), explained fire activity patterns at continental scales (Prior et al. 2017), and even allowed for interpretation of paleological reconstruction of fire activity (Belcher 2016). Furthermore, needle form and chemical characteristics in the foliage and litter bed have been employed to explain and develop fire spread and severity models (Schwilk and Caprio 2011; Torero and Simeoni 2010; Jolly et al. 2016; Grootemaat et al. 2017). Understanding leaf-level flammability has utilitarian applications in fire management.
Additional physical traits of jack pine trees should also be considered to explain their flammability at a landscape scale. Needle position, distribution and density in the twig and branches, needle age relative proportion, and crown architecture (e.g. height, bulk, and density) all modify the air/fuel mix in the crown and canopy, which has an important influence on landscape flammability (Rothermel 1972; Van Wagner 1977; Bond and Midgley 1995; Agee 1996; Schwilk 2003). For example, aggregated narrow, long needles provide high air circulation, enhancing their ignitability and combustibility, compared with shorter, wider needles that form more compact foliage (Scarff and Westoby 2006; Kane et al. 2008; Cornwell et al. 2015; Schwilk 2015). On the other hand, narrow needles spaced on the twigs spruce are highly ignitable, but the same needles generate a very compact fuel bed with low ignitable potential (Ganteaume 2018). More research is required at different spatial scales to discern the influence of chemical and physical traits on fire behaviour as additional tree features could enhance or counteract the effect of needle flammability
Conclusions
Far from behaving as a simple high moisture fuel, the flammability of living foliage is astoundingly intricate due to the complex relationships among the variables typical of living systems. We were able to experimentally determine the significance of living needle traits and their natural variation over the growing season on different flammable characteristics. Surprisingly, foliar moisture was not the primary driver of needle flammability, as is commonly assumed. Needle form significantly affected ignitability, whereas chemical composition affected combustibility and consumability. Our results suggest indirect effects of moisture content on flammability through form and chemical composition, but the nature of these interactions is still misunderstood. We advise studying vegetation fuels with an ecological approach, which will benefit wildland fire research and help us to understand the processes at finer spatial scales and improve fire behaviour modelling.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Support for this research came from the Canadian Partnership for Wildland Fire Science, Mike Flannigan NSERC Discovery Grant, CONACYT (scholarship 312318) and the Department of Renewable Resources at the University of Alberta.
Supplementary material
Supplementary material is available online.
Acknowledgements
We thank Dr. Philip G. Comeau for allowing us to work in his laboratory for the needle morphological analysis, Steven Paskaluk for the training and supervision on the calorimeter use and data retrieval, Dave Dunn for performing the chemical analysis, Dan Thompson for his advising on the testing methods, and Filmer Chu and Joseph Tse for their help editing this paper. We also thank students from the fire laboratory at the University of Alberta and friends for their support in collecting and preparing the samples: Xinli Cai, Luiz Drummond, Jorge Rodriguez, Rachel Keglowitsch, Chengtao Chen, Kimberly Morrison, Dante Castellanos, Jaime Sebastian, and Rafael Cabrera.
References
Agee JK (1996) The influence of forest structure on fire behavior. In ‘Proceedings of the 17th Annual Forest Vegetation Management Conference.’ 16–18 January 1996, Reeding, CA, USA. pp. 52–68. (University of California, Agriculture and Natural Resources: Berkeley, CA, USA)Alessio GA, Peñuelas J, Llusià J, Ogaya R, Estiarte M, de Lillis M (2008) Influence of water and terpenes on flammability in some dominant Mediterranean species. International Journal of Wildland Fire 17, 274–286.
| Influence of water and terpenes on flammability in some dominant Mediterranean species.Crossref | GoogleScholarGoogle Scholar |
Alexander ME, Cruz MG (2012) Interdependencies between flame length and fireline intensity in predicting crown fire initiation and crown scorch height. International Journal of Wildland Fire 21, 95–113.
| Interdependencies between flame length and fireline intensity in predicting crown fire initiation and crown scorch height.Crossref | GoogleScholarGoogle Scholar |
Alexander ME, Cruz MG (2013) Assessing the effect of foliar moisture on the spread rate of crown fires. International Journal of Wildland Fire 22, 415–427.
| Assessing the effect of foliar moisture on the spread rate of crown fires.Crossref | GoogleScholarGoogle Scholar |
Anderson HE (1970) Forest fuel ignitibility. Fire Technology 6, 312–319.
| Forest fuel ignitibility.Crossref | GoogleScholarGoogle Scholar |
Babrauskas V (1984) Development of the cone calorimeter – a bench‐scale heat release rate apparatus based on oxygen consumption. Fire and Materials 8, 81–95.
| Development of the cone calorimeter – a bench‐scale heat release rate apparatus based on oxygen consumption.Crossref | GoogleScholarGoogle Scholar |
Bakdash JZ, Marusich LR (2017) Repeated measures correlation. Frontiers in Psychology 8, 456
| Repeated measures correlation.Crossref | GoogleScholarGoogle Scholar |
Beckingham JD, Archibald JH (Eds) (1996) ‘Field Guide to Ecosites of Northern Alberta’,Vol. 5. (Natural Resources Canada, Canadian Forest Service, Northwest Region, Northern Forestry Centre: Edmonton, Alberta, Canada)
Behm AL, Duryea ML, Long AJ, Zipperer WC (2004) Flammability of native understory species in pine flatwood and hardwood hammock ecosystems and implications for the wildland–urban interface. International Journal of Wildland Fire 13, 355–365.
| Flammability of native understory species in pine flatwood and hardwood hammock ecosystems and implications for the wildland–urban interface.Crossref | GoogleScholarGoogle Scholar |
Belcher CM (2016) The influence of leaf morphology on litter flammability and its utility for interpreting palaeofire. Philosophical Transactions of the Royal Society B: Biological Sciences 371, 20150163
| The influence of leaf morphology on litter flammability and its utility for interpreting palaeofire.Crossref | GoogleScholarGoogle Scholar |
Bloom AJ, Chapin III FS, Mooney HA (1985) Resource limitation in plants – an economic analogy. Annual Review of Ecology and Systematics 16, 363–392.
| Resource limitation in plants – an economic analogy.Crossref | GoogleScholarGoogle Scholar |
Bond WJ, Midgley JJ (1995) Kill thy neighbour: an individualistic argument for the evolution of flammability. Oikos 73, 79
| Kill thy neighbour: an individualistic argument for the evolution of flammability.Crossref | GoogleScholarGoogle Scholar |
Bond WJ, van Wilgen BW (Eds) (1996) ‘Fire and Plants.’ (Springer Netherlands: Dordrecht, The Netherlands)
| Crossref |
Bond-Lamberty B, Wang C, Gower ST (2003) The use of multiple measurement techniques to refine estimates of conifer needle geometry. Canadian Journal of Forest Research 33, 101–105.
| The use of multiple measurement techniques to refine estimates of conifer needle geometry.Crossref | GoogleScholarGoogle Scholar |
Burgan RE (1979) Estimating live fuel moisture for the 1978 National Fire Danger Rating System. USDA Forest Service, Intermountain Forest and Range Experiment Station Paper INT–226. USDA Forest Service Ogden, UT, USA
Byram GM (1959) Combustion of forest fuels. In ‘Forest Fire Control and Use’. (Ed. K. P. Davis) pp. 61–89. (McGraw-Hill: Toronto, ON, Canada)
Campos-Ruiz R, Parisien M-A, Flannigan MD (2018) Temporal patterns of wildfire activity in areas of contrasting human influence in the Canadian boreal forest. Forests 9, 159
| Temporal patterns of wildfire activity in areas of contrasting human influence in the Canadian boreal forest.Crossref | GoogleScholarGoogle Scholar |
Carson P, Mumford C (Eds) (1994) ‘Hazardous Chemicals Handbook.’ (Butterworth–Heinemann: Oxford, UK)
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18, 117–143.
| Non-parametric multivariate analyses of changes in community structure.Crossref | GoogleScholarGoogle Scholar |
Cornwell WK, Elvira A, van Kempen L, van Logtestijn RSP, Aptroot A, Cornelissen JHC (2015) Flammability across the gymnosperm phylogeny: the importance of litter particle size. New Phytologist 206, 672–681.
| Flammability across the gymnosperm phylogeny: the importance of litter particle size.Crossref | GoogleScholarGoogle Scholar |
Cruz MG, Alexander ME, Wakimoto RH (2005) Development and testing of models for predicting crown fire rate of spread in conifer forest stands. Canadian Journal of Forest Research 35, 1626–1639.
| Development and testing of models for predicting crown fire rate of spread in conifer forest stands.Crossref | GoogleScholarGoogle Scholar |
Darwish Ahmad A, Abubaker AM, Salaimeh A, Akafuah NK, Finney M, Forthofer JM, Saito K (2021) Ignition and burning mechanisms of live spruce needles. Fuel 304, 121371
| Ignition and burning mechanisms of live spruce needles.Crossref | GoogleScholarGoogle Scholar |
de Groot WJ, Flannigan MD, Cantin AS (2013) Climate change impacts on future boreal fire regimes. Forest Ecology and Management 294, 35–44.
| Climate change impacts on future boreal fire regimes.Crossref | GoogleScholarGoogle Scholar |
de Lillis M, Bianco PM, Loreto F (2009) The influence of leaf water content and isoprenoids on flammability of some Mediterranean woody species. International Journal of Wildland Fire 18, 203–212.
| The influence of leaf water content and isoprenoids on flammability of some Mediterranean woody species.Crossref | GoogleScholarGoogle Scholar |
Della Rocca G, Madrigal J, Marchi E, Michelozzi M, Moya B, Danti R (2017) Relevance of terpenoids on flammability of Mediterranean species: an experimental approach at a low radiant heat flux. IForest - Biogeosciences and Forestry 10, 766–775.
| Relevance of terpenoids on flammability of Mediterranean species: an experimental approach at a low radiant heat flux.Crossref | GoogleScholarGoogle Scholar |
Della Rocca G, Danti R, Hernando C, Guijarro M, Michelozzi M, Carrillo C, Madrigal J (2020) Terpenoid accumulation links plant health and flammability in the cypress-bark canker pathosystem. Forests 11, 651
| Terpenoid accumulation links plant health and flammability in the cypress-bark canker pathosystem.Crossref | GoogleScholarGoogle Scholar |
Dewhirst RA, Smirnoff N, Belcher CM (2020) Pine species that support crown fire regimes have lower leaf-level terpene contents than those native to surface fire regimes. Fire 3, 17
| Pine species that support crown fire regimes have lower leaf-level terpene contents than those native to surface fire regimes.Crossref | GoogleScholarGoogle Scholar |
Dimitrakopoulos AP, Papaioannou KK (2001) Flammability assessment of Mediterranean forest fuels. Fire Technology 37, 143–152.
| Flammability assessment of Mediterranean forest fuels.Crossref | GoogleScholarGoogle Scholar |
Etlinger MG, Beall FC (2004) Development of a laboratory protocol for fire performance of landscape plants. International Journal of Wildland Fire 13, 479–488.
| Development of a laboratory protocol for fire performance of landscape plants.Crossref | GoogleScholarGoogle Scholar |
Farrar JL (Ed.) (1995) ‘Trees of the Northern United States and Canada.’ (Iowa State University Press: Ames, IA, USA)
Faul F, Erdfelder E, Lang A-G, Buchner A (2007) G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods 39, 175–191.
| G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences.Crossref | GoogleScholarGoogle Scholar |
Fazeli H, Jolly WM, Blunck DL (2022) Stages and time-scales of ignition and burning of live fuels for different convective heat fluxes. Fuel 324, 124490
| Stages and time-scales of ignition and burning of live fuels for different convective heat fluxes.Crossref | GoogleScholarGoogle Scholar |
Fernandes PM, Cruz MG (2012) Plant flammability experiments offer limited insight into vegetation-fire dynamics interactions. New Phytologist 194, 606–609.
| Plant flammability experiments offer limited insight into vegetation-fire dynamics interactions.Crossref | GoogleScholarGoogle Scholar |
Finney MA, Cohen JD, McAllister SS, Jolly WM (2013) On the need for a theory of wildland fire spread. International Journal of Wildland Fire 22, 25–36.
| On the need for a theory of wildland fire spread.Crossref | GoogleScholarGoogle Scholar |
Fischer C, Höll W (1991) Food reserves of Scots pine (Pinus sylvestris L.). Trees 5, 187–195.
| Food reserves of Scots pine (Pinus sylvestris L.).Crossref | GoogleScholarGoogle Scholar |
Fletcher TH, Pickett BM, Smith SG, Spittle GS, Woodhouse MM, Haake E, Weise DR (2007) Effects of moisture on ignition behavior of moist California Chaparral and Utah leaves. Combustion Science and Technology 179, 1183–1203.
| Effects of moisture on ignition behavior of moist California Chaparral and Utah leaves.Crossref | GoogleScholarGoogle Scholar |
Friedrich S, Konietschke F, Pauly M (2018) Analysis of multivariate data and repeated measures designs with the R Package MANOVA.RM. arXiv preprint arXiv:1801.08002
Ganteaume A (2018) Does plant flammability differ between leaf and litter bed scale? Role of fuel characteristics and consequences for flammability assessment. International Journal of Wildland Fire 27, 342–352.
| Does plant flammability differ between leaf and litter bed scale? Role of fuel characteristics and consequences for flammability assessment.Crossref | GoogleScholarGoogle Scholar |
Ganteaume A, Romero B, Fernandez C, Ormeño E, Lecareux C (2021) Volatile and semi-volatile terpenes impact leaf flammability: differences according to the level of terpene identification. Chemoecology 31, 259–275.
| Volatile and semi-volatile terpenes impact leaf flammability: differences according to the level of terpene identification.Crossref | GoogleScholarGoogle Scholar |
Gauthier S, Bergeron Y, Simon J-P (1993) Cone serotiny in jack pine: ontogenetic, positional, and environmental effects. Canadian Journal of Forest Research 23, 394–401.
| Cone serotiny in jack pine: ontogenetic, positional, and environmental effects.Crossref | GoogleScholarGoogle Scholar |
Goslee SC, Urban DL (2007) The ecodist package for dissimilarity-based analysis of ecological data. Journal of Statistical Software 22, 1–19.
| The ecodist package for dissimilarity-based analysis of ecological data.Crossref | GoogleScholarGoogle Scholar |
Government of Canada (2019) Environmental and natural resources: monthly climate summaries. Available at https://climate.weather.gc.ca/prods_servs/cdn_climate_summary_e.html [Verified 9 September 2019]
Greenberg JP, Friedli H, Guenther AB, Hanson D, Harley P, Karl T (2006) Volatile organic emissions from the distillation and pyrolysis of vegetation. Atmospheric Chemistry and Physics 6, 81–91.
| Volatile organic emissions from the distillation and pyrolysis of vegetation.Crossref | GoogleScholarGoogle Scholar |
Grootemaat S, Wright IJ, van Bodegom PM, Cornelissen JHC (2017) Scaling up flammability from individual leaves to fuel beds. Oikos 126, 1428–1438.
| Scaling up flammability from individual leaves to fuel beds.Crossref | GoogleScholarGoogle Scholar |
Guerrero F, Hernández C, Toledo M, Espinoza L, Carrasco Y, Arriagada A, Muñoz A, Taborga L, Bergmann J, Carmona C (2021) Leaf thermal and chemical properties as natural drivers of plant flammability of native and exotic tree species of the Valparaíso region, Chile. International Journal of Environmental Research and Public Health 18, 7191
| Leaf thermal and chemical properties as natural drivers of plant flammability of native and exotic tree species of the Valparaíso region, Chile.Crossref | GoogleScholarGoogle Scholar |
Hawley LF (1926) Theoretical considerations regarding factors which influence forest fires. Journal of Forestry 24, 756–763.
| Theoretical considerations regarding factors which influence forest fires.Crossref | GoogleScholarGoogle Scholar |
Heinselman ML (1973) Fire in the virgin forests of the Boundary Waters Canoe Area, Minnesota. Quaternary Research 3, 329–382.
| Fire in the virgin forests of the Boundary Waters Canoe Area, Minnesota.Crossref | GoogleScholarGoogle Scholar |
Hoch G, Richter A, Körner C (2003) Non-structural carbon compounds in temperate forest trees. Plant, Cell and Environment 26, 1067–1081.
| Non-structural carbon compounds in temperate forest trees.Crossref | GoogleScholarGoogle Scholar |
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biometrical Journal 50, 346–363.
| Simultaneous inference in general parametric models.Crossref | GoogleScholarGoogle Scholar |
Hubert EE (1932) Briefer articles and notes: A brief study of conifer needle oils. Journal of Forestry 30, 340–344.
| Briefer articles and notes: A brief study of conifer needle oils.Crossref | GoogleScholarGoogle Scholar |
Jervis FX, Rein G (2016) Experimental study on the burning behaviour of Pinus halepensis needles using small-scale fire calorimetry of live, aged and dead samples. Fire and Materials 40, 385–395.
| Experimental study on the burning behaviour of Pinus halepensis needles using small-scale fire calorimetry of live, aged and dead samples.Crossref | GoogleScholarGoogle Scholar |
Johnson VJ (1966) ‘Seasonal Fluctuation in Moisture Content of Pine Foliage.’ (USDA North Central Forest Experiment Station: St. Paul, MN, USA)
Jolly WM, Parsons RA, Hadlow AM, Cohn GM, McAllister SS, Popp JB, Hubbard RM, Negron JF (2012) Relationships between moisture, chemistry, and ignition of Pinus contorta needles during the early stages of mountain pine beetle attack. Forest Ecology and Management 269, 52–59.
| Relationships between moisture, chemistry, and ignition of Pinus contorta needles during the early stages of mountain pine beetle attack.Crossref | GoogleScholarGoogle Scholar |
Jolly WM, Hintz J, Linn RL, Kropp RC, Conrad ET, Parsons RA, Winterkamp J (2016) Seasonal variations in red pine (Pinus resinosa) and jack pine (Pinus banksiana) foliar physio-chemistry and their potential influence on stand-scale wildland fire behavior. Forest Ecology and Management 373, 167–178.
| Seasonal variations in red pine (Pinus resinosa) and jack pine (Pinus banksiana) foliar physio-chemistry and their potential influence on stand-scale wildland fire behavior.Crossref | GoogleScholarGoogle Scholar |
Kane JM, Varner JM, Hiers JK (2008) The burning characteristics of southeastern oaks: Discriminating fire facilitators from fire impeders. Forest Ecology and Management 256, 2039–2045.
| The burning characteristics of southeastern oaks: Discriminating fire facilitators from fire impeders.Crossref | GoogleScholarGoogle Scholar |
Kanury AM (1994) Combustion characteristics of biomass fuels. Combustion Science and Technology 97, 469–491.
| Combustion characteristics of biomass fuels.Crossref | GoogleScholarGoogle Scholar |
Kozlowski TT, Kramer PJ, Pallardy SG (1991) Physiological and environmental requirements for tree growth. In ‘The Physiological Ecology of Woody Plants’. (Eds TT Kozlowski, PJ Kramer, SG Pallardy) pp. 31–68. (Elsevier: San Diego, CA, USA)
| Crossref |
Kramer PJ, Kozlowski TT (Eds) (1979) ‘Physiology of Woody Plants.’ (Academic Press: New York, USA)
| Crossref |
Krix DW, Phillips ML, Murray BR (2019) Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland–urban interface. International Journal of Wildland Fire 28, 295–307.
| Relationships among leaf flammability attributes and identifying low-leaf-flammability species at the wildland–urban interface.Crossref | GoogleScholarGoogle Scholar |
Langenheim JH (1994) Higher plant terpenoids: a phytocentric overview of their ecological roles. Journal of Chemical Ecology 20, 1223–1280.
| Higher plant terpenoids: a phytocentric overview of their ecological roles.Crossref | GoogleScholarGoogle Scholar |
Lenth RV (2016) Least-squares means: the R package lsmeans. Journal of Statistical Software 69, 1–33.
| Least-squares means: the R package lsmeans.Crossref | GoogleScholarGoogle Scholar |
Little CHA (1970) Seasonal changes in carbohydrate and moisture content in needles of balsam fir (Abies balsamea). Canadian Journal of Botany 48, 2021–2028.
| Seasonal changes in carbohydrate and moisture content in needles of balsam fir (Abies balsamea).Crossref | GoogleScholarGoogle Scholar |
Little EL (1979) ‘Checklist of United States trees (native and naturalized).’ (Forest Service, US Department of Agriculture: Washington, DC, USA)
Mandre M, Tullus H, Klõšeiko J (2002) Partitioning of carbohydrates and biomass of needles in Scots pine canopy. Zeitschrift fur Naturforschung - Section C Journal of Biosciences 57, 296–302.
| Partitioning of carbohydrates and biomass of needles in Scots pine canopy.Crossref | GoogleScholarGoogle Scholar |
Martin RE, Gordon DA, Gutierrez ME, Lee DS, Molina DM, Schroeder RA, Sapsis DB, Stephens SL, Chambers M (1994) Assessing the flammability of domestic and wildland vegetation. In ‘12th Conference on Fire and Forest Meteorology.’ 26–28 May, Jekyll Island, GA, USA. pp. 130–137. (Society of American Foresters: Bathesda, MD, USA)
McAllister S, Grenfell I, Hadlow A, Jolly WM, Finney M, Cohen J (2012) Piloted ignition of live forest fuels. Fire Safety Journal 51, 133–142.
| Piloted ignition of live forest fuels.Crossref | GoogleScholarGoogle Scholar |
Melnik OM, Paskaluk SA, Ackerman MY, Melnik KO, Thompson DK, McAllister SS, Flannigan MD (2022) New in-flame flammability testing method applied to monitor seasonal changes in live fuel. Fire 5, 1
| New in-flame flammability testing method applied to monitor seasonal changes in live fuel.Crossref | GoogleScholarGoogle Scholar |
Montgomery KR, Cheo PC (1971) Effect of leaf thickness on ignitibility. Forest Science 17, 475–478.
| Effect of leaf thickness on ignitibility.Crossref | GoogleScholarGoogle Scholar |
Murray BR, Hardstaff LK, Phillips ML (2013) Differences in leaf flammability, leaf traits and flammability-trait relationships between native and exotic plant species of dry sclerophyll forest. PloS One 8, e79205
| Differences in leaf flammability, leaf traits and flammability-trait relationships between native and exotic plant species of dry sclerophyll forest.Crossref | GoogleScholarGoogle Scholar |
Murray BR, Brown C, Murray ML, Krix DW, Martin LJ, Hawthorne T, Wallace MI, Potvin SA, Webb JK (2020) An integrated approach to identify low-flammability plant species for green firebreaks. Fire 3, 9
| An integrated approach to identify low-flammability plant species for green firebreaks.Crossref | GoogleScholarGoogle Scholar |
Naresh K, Kumar A, Korobeinichev O, Shmakov A, Osipova K (2018) Downward flame spread along a single pine needle: numerical modelling. Combustion and Flame 197, 161–181.
| Downward flame spread along a single pine needle: numerical modelling.Crossref | GoogleScholarGoogle Scholar |
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Henry M, Stevens MHH, Szoecs E, Wagner H (2020) vegan: community ecology package. R package version 2.5–7. Available at http://CRAN.R-project.org/package=vegan
Owens MK, Lin C-D, Taylor Jr CA, Whisenant SG (1998) Seasonal patterns of plant flammability and monoterpenoid content in Juniperus ashei. Journal of Chemical Ecology 24, 2115–2129.
| Seasonal patterns of plant flammability and monoterpenoid content in Juniperus ashei.Crossref | GoogleScholarGoogle Scholar |
Page WG, Jenkins MJ, Runyon JB (2012) Mountain pine beetle attack alters the chemistry and flammability of lodgepole pine foliage. Canadian Journal of Forest Research 42, 1631–1647.
| Mountain pine beetle attack alters the chemistry and flammability of lodgepole pine foliage.Crossref | GoogleScholarGoogle Scholar |
Papió C, Trabaud L (1990) Structural characteristics of fuel components of five Mediterranean shrubs. Forest Ecology and Management 35, 249–259.
| Structural characteristics of fuel components of five Mediterranean shrubs.Crossref | GoogleScholarGoogle Scholar |
Patnaik P (Ed.) (2007) ‘A Comprehensive Guide to the Hazardous Properties of Chemical Substances.’ (Wiley & Sons: Hoboken, NJ, USA)
Pausas JG, Alessio GA, Moreira B, Segarra-Moragues JG (2016) Secondary compounds enhance flammability in a Mediterranean plant. Oecologia 180, 103–110.
| Secondary compounds enhance flammability in a Mediterranean plant.Crossref | GoogleScholarGoogle Scholar |
Philpot C (1970) Influence of mineral content on the pyrolysis of plant materials. Forest Science 16, 46–471.
Philpot CW, Mutch RW (1971) The seasonal trends in moisture content, ether extractives, and energy of ponderosa pine and douglas fir needles. Intermountain Forest and Range Experiment Station Research Paper INT-102, USDA Forest Service, Ogden, UT, USA
Pickett BM, Isackson C, Wunder R, Fletcher TH, Butler BW, Weise DR (2010) Experimental measurements during combustion of moist individual foliage samples. International Journal of Wildland Fire 19, 153–162.
| Experimental measurements during combustion of moist individual foliage samples.Crossref | GoogleScholarGoogle Scholar |
Popović Z, Bojović S, Marković M, Cerdà A (2021) Tree species flammability based on plant traits: a synthesis. Science of the Total Environment 800, 149625
| Tree species flammability based on plant traits: a synthesis.Crossref | GoogleScholarGoogle Scholar |
Prior LD, Murphy BP, Williamson GJ, Cochrane MA, Jolly WM, Bowman DMJS (2017) Does inherent flammability of grass and litter fuels contribute to continental patterns of landscape fire activity. Journal of Biogeography 44, 1225–1238.
| Does inherent flammability of grass and litter fuels contribute to continental patterns of landscape fire activity.Crossref | GoogleScholarGoogle Scholar |
Pyne SJ (2007) Problems, paradoxes, paradigms: triangulating fire research. International Journal of Wildland Fire 16, 271–276.
| Problems, paradoxes, paradigms: triangulating fire research.Crossref | GoogleScholarGoogle Scholar |
Pyne S, Andrews P, Laven R (Eds) (1996) ‘Introduction to Wildland Fire.’ (Wiley: New York, USA)
R Core Team (2008) R: A language and environment for statistical computing. Available at http://www.r-project.org/
Raman V, Sivasankaralingam V, Dibble R, Sarathy S (2016) α-Pinene – a high energy density biofuel for SI engine applications. SAE Technical Paper 2016-01-2171
| α-Pinene – a high energy density biofuel for SI engine applications.Crossref | GoogleScholarGoogle Scholar |
Raven JA (1998) Plant Resource Allocation. In ‘Trends in Ecology & Evolution’. (Eds FA Bazzaz, J Grace) Vol. 13, pp. 84–85.
| Crossref |
Rivera JD, Davies GM, Jahn W (2012) Flammability and the heat of combustion of natural fuels: a review. Combustion Science and Technology 184, 224–242.
| Flammability and the heat of combustion of natural fuels: a review.Crossref | GoogleScholarGoogle Scholar |
Romero B, Fernandez C, Lecareux C, Ormeño E, Ganteaume A (2019) How terpene content affects fuel flammability of wildland–urban interface vegetation. International Journal of Wildland Fire 28, 614–627.
| How terpene content affects fuel flammability of wildland–urban interface vegetation.Crossref | GoogleScholarGoogle Scholar |
Rossa CG, Fernandes PM (2018) Live fuel moisture content: The ‘pea under the mattress’ of fire spread rate modeling? Fire 1, 43
| Live fuel moisture content: The ‘pea under the mattress’ of fire spread rate modeling?Crossref | GoogleScholarGoogle Scholar |
Rothermel RC (1972) A mathematical model for predicting fire spread in wildland fuels. Intermountain Forest and Range Experiment Station Research Paper INT-115, USDA Forest Service: Ogden, UT, USA
Scarff FR, Westoby M (2006) Leaf litter flammability in some semi-arid Australian woodlands. Functional Ecology 20, 745–752.
| Leaf litter flammability in some semi-arid Australian woodlands.Crossref | GoogleScholarGoogle Scholar |
Schoonmaker A (2013) Resource Allocation, Water Relations and Crown Architecture Examined at the Tree and Stand-level in Northern Conifers. PhD thesis, University of Alberta, Edmonton.
Schwilk DW (2003) Flammability is a niche construction trait: canopy architecture affects fire intensity. The American naturalist 162, 725–33.
| Flammability is a niche construction trait: canopy architecture affects fire intensity.Crossref | GoogleScholarGoogle Scholar |
Schwilk DW (2015) Dimensions of plant flammability. New Phytologist 206, 486–488.
| Dimensions of plant flammability.Crossref | GoogleScholarGoogle Scholar |
Schwilk DW, Caprio AC (2011) Scaling from leaf traits to fire behaviour: community composition predicts fire severity in a temperate forest. Journal of Ecology 99, 970–980.
| Scaling from leaf traits to fire behaviour: community composition predicts fire severity in a temperate forest.Crossref | GoogleScholarGoogle Scholar |
Shen C, Fletcher TH (2015) Fuel element combustion properties for live wildland Utah shrubs. Combustion Science and Technology 187, 428–444.
| Fuel element combustion properties for live wildland Utah shrubs.Crossref | GoogleScholarGoogle Scholar |
Simeoni A, Thomas JC, Bartoli P, Borowieck P, Reszka P, Colella F, Santoni PA, Torero JL (2012) Flammability studies for wildland and wildland–urban interface fires applied to pine needles and solid polymers. Fire Safety Journal 54, 203–217.
| Flammability studies for wildland and wildland–urban interface fires applied to pine needles and solid polymers.Crossref | GoogleScholarGoogle Scholar |
Simms DL, Law M (1967) The ignition of wet and dry wood by radiation. Combustion and Flame 11, 377–388.
| The ignition of wet and dry wood by radiation.Crossref | GoogleScholarGoogle Scholar |
Torero JL, Simeoni A (2010) Heat and mass transfer in fires: scaling laws, ignition of solid fuels and application to forest fires. The Open Thermodynamics Journal 4, 145–155.
| Heat and mass transfer in fires: scaling laws, ignition of solid fuels and application to forest fires.Crossref | GoogleScholarGoogle Scholar |
Van Wagner CE (1977) Conditions for the start and spread of crown fire. Canadian Journal of Forest Research 7, 23–34.
| Conditions for the start and spread of crown fire.Crossref | GoogleScholarGoogle Scholar |
Van Wagner CE (1987) Development and structure of the Canadian Forest Fire Weather Index System. Canadian Forestry Service Forestry Technical Report 35, Canadian Forestry Service: Ottawa, ON, Canada
Van Wagner CE (1998) Modelling logic and the Canadian Forest Fire Behavior Prediction System. The Forestry Chronicle 74, 50–52.
| Modelling logic and the Canadian Forest Fire Behavior Prediction System.Crossref | GoogleScholarGoogle Scholar |
van Wilgen BW, Higgins KB, Bellstedt DU (1990) The role of vegetation structure and fuel chemistry in excluding fire from forest patches in the fire-prone fynbos shrublands of South Africa. Journal of Ecology 78, 210–222.
| The role of vegetation structure and fuel chemistry in excluding fire from forest patches in the fire-prone fynbos shrublands of South Africa.Crossref | GoogleScholarGoogle Scholar |
Virjamo V, Julkunen-Tiitto R (2016) Pinaceae alkaloids. In ‘Plant Specialized Metabolism’. (Eds G Arimura, M Maffei) pp. 131–142. (CRC Press: New York, USA)
Vose JM, Ryan MG (2002) Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis. Global Change Biology 8, 182–193.
| Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis.Crossref | GoogleScholarGoogle Scholar |
Weise DR, White RH, Beall FC, Etlinger M (2005) Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation. International Journal of Wildland Fire 14, 321–338.
| Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation.Crossref | GoogleScholarGoogle Scholar |
White R (1987) Effect of lignin content and extractives on the higher heating value of wood. Wood and Fiber Science 19, 446–452.
White RH, Zipperer WC (2010) Testing and classification of individual plants for fire behaviour: plant selection for the wildland–urban interface. International Journal of Wildland Fire 19, 213–227.
| Testing and classification of individual plants for fire behaviour: plant selection for the wildland–urban interface.Crossref | GoogleScholarGoogle Scholar |
Zakzeski J, Bruijnincx PCA, Jongerius AL, Weckhuysen BM (2010) The catalytic valorization of lignin for the production of renewable chemicals. Chemical Reviews 110, 3552–3599.
| The catalytic valorization of lignin for the production of renewable chemicals.Crossref | GoogleScholarGoogle Scholar |