

Virtual phyllotaxis and real plant model cases
Beata Zagórska-Marek A B and Marcin Szpak AA Institute of Plant Biology, Wrocław University, 50-328 Wrocław, Kanonia Street 6/8, Poland.
B Corresponding author. Email: beata@biol.uni.wroc.pl
This paper originates from a presentation at the 5th International Workshop on Functional–Structural Plant Models, Napier, New Zealand, November 2007.
Functional Plant Biology 35(10) 1025-1033 https://doi.org/10.1071/FP08076
Submitted: 14 March 2008 Accepted: 4 August 2008 Published: 11 November 2008
Abstract
Phyllotactic pattern results from genetic control of lateral primordia size (physiological or physical) relative to the size of organogenic lateral surface of shoot apical meristem (SAM). In order to understand the diversity of patterns and ontogenetic transitions of phyllotaxis we have developed a geometric model allowing changes of the above proportion in a computer simulation of SAM’s growth. The results of serial simulations confirmed that many phyllotactic patterns (including most esoteric ones) and ontogenetic transitions known from real plant model cases can be easily obtained in silico. Properties of virtual patterns often deviated from those of ideal mathematical lattices but closely resembled those of the natural ones. This proved the assumptions of the model, such as initiation in the first available space or ontogenetic changes in primordia size, to be quite realistic. Confrontation of simulation results with some sequences of real phyllotactic patterns (case study Verbena) questions the autonomy of SAM in its organogenic activity and suggests the involvement of unknown signal positioning primordia in a non-random manner in the first available space.
Additional keywords: Magnolia, ontogenetic transitions, pattern formation, SAM, Verbena.
MAGNOLIA
Sleepy flower on the leaf
Creamy-white ivory
Sweet to dizziness
Fragrant thing
Maliciously mysterious Universe
Strange visitor among us, humans
(Maria Pawlikowska-Jasnorzewska 1924)
Introduction
Phyllotaxis is the distribution of lateral organs in plants. The organ primordia are initiated iteratively (Hofmeister 1868) on the organogenic surface of the shoot apical meristem (SAM). The regularity of phyllotactic patterns has always intrigued scientists. Today there is still no exaggeration in quoting Darwin’s famous statement and saying, that both phyllotactic diversity and, especially, ontogenetic transitions of phyllotaxis remain an ‘abominable mystery’ in plant biology. The phylogenetic aspect of the diversity is that phyllotactic patterns can be species specific. Having the status of a diagnostic feature in plant taxonomy, they fall into two major categories: whorled – achiral patterns, and helical – chiral patterns. Within one category there are many types of patterns: the decussate or tricussate in the first one, the main Fibonacci, Lucas and many others in the second. The ontogenetic aspect of phyllotactic diversity is that the pattern often changes together with the developmental phase of plant’s growth. From the biological point of view it is, however, the ontogenetic, qualitative changes of phyllotaxis, which occur for no apparent reason and are not associated with the developmental switch of plant organ identity, that are truly challenging. The ontogenetic transitions between the patterns are qualitative when the category or the type of pattern changes, or quantitative when, especially in chiral patterns, there is a progressive change in phyllotactic order (expression) i.e. in the numbers of contact parastichies (Adler 1974, 1977).
The mechanism positioning the primordia of leaves, flowers or flower parts on the organogenic surface of SAM has not yet been fully elucidated. It is undoubtedly genetically controlled (phylogenetic aspect) but also flexible enough to assure the plant’s phenotypic plasticity (ontogenetic transitions).
The latest developments in plant biology point out the role of auxin, transported acropetally, in the superficial cellular layer of SAM (Reinhardt et al. 2000, 2003; Heisler et al. 2005). The polarity of the cells transporting the hormone alters periodically in the vicinity of emerging primordium, with the flux of auxin being once oriented towards the primordium tip and, after a while, away from it, towards the tip of the apex itself. A spacing mechanism is created by the feedback of the hormone on its own transport. Its component is a primordium acting as a sink, competing with others for the hormone and inhibiting the formation of similar structures in its close vicinity. The efficiency of a sink can be measured by the size of the inhibition field, within which, the auxin concentration is below the level required for the formation of a new sink. New elements emerge in the first available space between already existing neighbouring primordia, as postulated earlier by Snow and Snow (1931, 1952). This space is created by the constant and continuous addition of new cells in a process of SAM’s apical growth. Based on these concepts two new models of phyllotaxis have recently been introduced (Jönsson et al. 2006; Smith et al. 2006a). We still do not know though what kind of signal changes the polarity of the cells and reorients the flux of auxin. Moreover, it is still uncertain to what degree the SAM is autonomous in positioning organ primordia and what role the signals, which flow acropetally from differentiated tissues play in this process (Larson 1977; Banasiak and Zagórska-Marek 2006).
Phyllotactic patterns result from genetic control of lateral primordia size (physiological or physical) relative to the size of organogenic lateral surface of the SAM. It is reasonable to assume that the change in one of these parameters alters the ratio and thus brings up a change in phyllotaxis. The aim of this study was to find, in a computer simulation, the causes of phyllotactic transitions known from plant model cases. The principles affecting the direction of phyllotactic transitions and, thus, the quality of the emerging patterns were of particular interest.
The model
The physiological size of primordia may have nothing to do with their real geometric size. However, in the models of phyllotactic pattern formation and the pattern’s subsequent transformation, it is convenient to have the primordia (or their inhibition fields) shown as circles tightly packed on a circular, cylindrical, paraboloid or conic surface representing organogenic surface of growing SAM (Williams and Brittain 1984; Battjes and Prusinkiewicz 1998; Zagórska-Marek 1999, 2000; Smith et al. 2006b; Hellwig et al. 2008).
In order to understand the emergence of various patterns and their ontogenetic transitions, we have simulated SAM’s growth using a special computer program written by Marcin Szpak (Zagórska-Marek and Szpak 2006). In our geometric model we assumed:
-
an infinite cylinder of a constant width as simulation space,
-
primordia as circles of a changing radius, and
-
primordia emerging in the first available space (in lowest possible position between the two neighbouring, already existing primordia).
The following abbreviations of simulation parameters have been used throughout the paper.
-
R, the radius of the circle representing organ primordium,
-
IP, the initial phyllotactic pattern,
-
PA, the angle of parastichy intersection in the initial phyllotactic pattern,
-
DR, the deviation rate, i.e. the change in the radius of subsequent primordium, expressed as percentage of the previous value of R (in case of continuous change in R),
-
DS, the percentage change (in case of discontinuous change in R), and
-
DM, the deviation minimum, i.e. the limit of the change in R, expressed as percentage of the initial value of R.
In interpreting the results of simulations, we have identified the emerging phyllotactic pattern by counting its contact parastichies and writing down their numbers in the opposed parastichy pair formula (Adler 1974, 1977). The numbers were supplemented with s and z indexes, which allowed identification of pattern chirality. s was used when orientation of parastichies on the organogenic surface viewed from the outside was to the left, z, when it was to the right. Besides the contact parastichies tangent to each other, thus, being the most conspicuous, there are also many other secondary helices in each phyllotactic pattern. Their numbers belong to the mathematical series associated with the pattern (as the 1, 2, 3, 5, 8 … series with the main Fibonacci pattern). They are inconspicuous but may become visible when the order (expression) of phyllotaxis changes. Their numbers and orientation are also informative, helpful in pattern identification. The list of patterns identified in nature and of the mathematical series attributed to them (based on secondary connections between the pattern elements) has been provided in earlier papers (Zagórska-Marek 1985, 1994).
Results and discussion
The assumption that the size of primordia is subject to ontogenetic change appeared to be particularly fruitful. The model, in testing, produced an almost infinite number of transitions, with many resulting patterns of primordia spacing, more or less regular. The changes we had been applying were abrupt or continuous, quick or slow, with the radius decreasing or increasing.
Continuous change
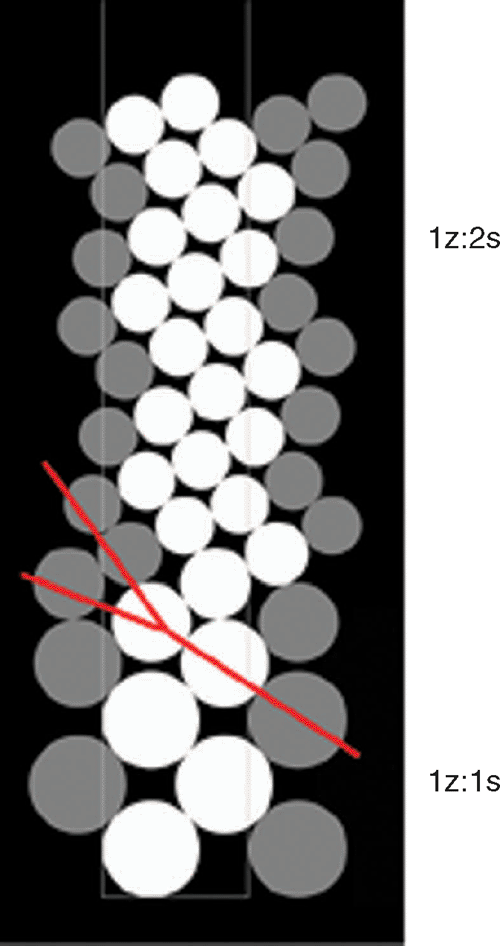
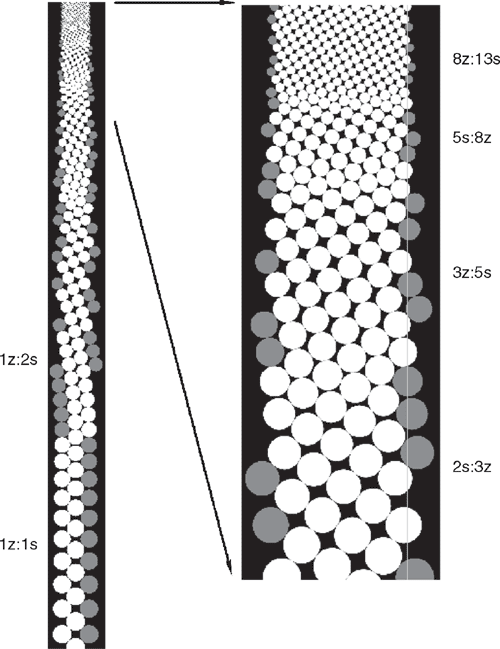
The continuous decrease or increase in the radius of pattern elements (R) typically changed an order (expression) of the initial phyllotactic pattern, which is quite a common condition in many plants. We have investigated this case assuming the initial pattern (IP) as distichous, with the expression 1 : 1. This pattern, the first in monocots, where there is only one cotyledon in the embryo, often extends to the vegetative zone of the main axis, as exemplified by grasses. In dicots, as they possess two cotyledons, the IP should be decussate (see the case study Verbena below). In the simulation the continuous decrease in R (Fig. 1), caused transformation of the initial 1 : 1 distichous pattern into 1 : 2 main Fibonacci phyllotaxis. The decrease systematically widened the available space, to the point when new primordium could be shifted in it either to the left or to the right. It was a moment of symmetry breaking. Random selection of the shift set up the chirality of the emerging pattern, but it did not affect its type – in both situations it was the main Fibonacci pattern. This qualitative transition was accomplished through dislocation (red line on Fig. 1), i.e. by an addition of one parastichy in only one of the two sets of helices making the opposed parastichy pair (sensu Adler 1974, 1977). Similar dislocations, associated with qualitative transitions between various patterns, have been described in natural phyllotactic lattices of balsam fir and magnolia (Zagórska-Marek 1985, 1987, 1994). In a simulation, further changes in R caused the continuous rise in order of the chiral pattern (Fig. 2). This case is best illustrated in nature by inflorescences of Asteraceae family. In capitula of Helianthus, Bellis or Telekia (Fig. 3), the numbers of parastichies in the opposed parastichy pair are usually very high. They represent one of the highest expressions of main Fibonacci phyllotaxis known in plant kingdom. Vegetative phyllotaxis in Asteraceae, preceding the formation of inflorescence, is of a much lower order (Couder 1998). Tracing selected parastichies during these ontogenetic changes shows that they visibly change their inclination, and ultimately become inconspicuous. We have observed the same effect in the simulation where IP was the main Fibonacci (Fig. 4) and R was continuously increasing. In this case, the order of phyllotaxis was decreasing. In all our simulations, the width of the organogenic surface was constant. In nature, the changes in order of phyllotaxis are often observed when pattern elements are of the same identity – in coniferous vegetative shoots or in some inflorescences (Fig. 4). It is likely that in these cases they are caused by developmental fluctuations in a size of SAM’s organogenic surface, associated with changing vigor of the shoot, or with determinate character of its growth (as in capitula of Asteraceae).
|
|

|

|
We noted a dependence of the quality of the emerging pattern on the initial parastichy angle (PA). We have tested the significance of PA in a sequence of simulations by changing its value from 60 to 90°. Other parameters of the simulations were kept constant at the following values: IP, 1 : 1; DR, 97%; DM, 40%. The emerging pattern was either the esoteric 1, 2, 3, 8, 11… phyllotaxis (60 < PA >78.8) or the common 1, 2, 3, 5, 8… main Fibonacci pattern (PA >78.8). In both these patterns, shown in Fig. 5, the orientation of low order parastichies was identical: 2s : 3z, but 8-parastichies (red asterisks) in a resulting pattern, initially oriented to the left (s), became vertical when PA = 78.8 degrees and then flipped to the right (z). The low 2 : 3 expression causes that the qualitatively different pattern shown in the left simulation in Fig. 5 may be erroneously classified as the main Fibonacci. Its divergence, however, is 129.76, close to the limiting value. The latter, calculated theoretically, equals 132.18, whereas for the main Fibonacci – 137.5. We note that the first pattern, the appearance of which in computer simulations was also reported by Liaw (1998), belongs to the so-called ‘impossible’ patterns (Jean 1994). It has been reported by Fujita (1937, 1939) as appearing sporadically in Plantago, by Zagórska-Marek (1985) as engaged in phyllotactic transitions in Abies balsamea vegetative shoots and finally it was documented on scanning electron microphotographs of Magnolia floral apices (Zagórska-Marek 1994).

|
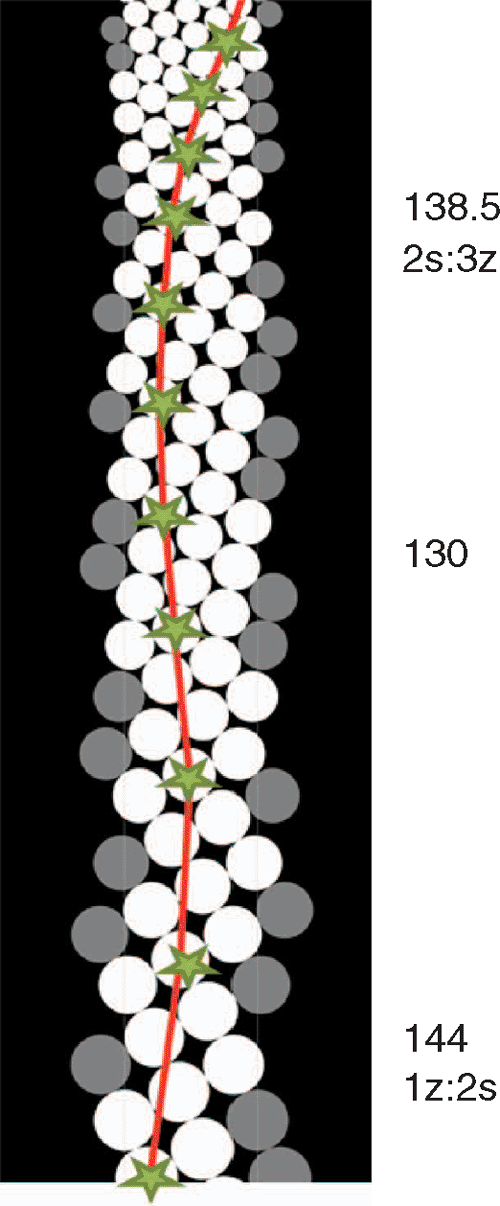
Intrigued by the appearance of this pattern in our simulations, we traced the consecutive divergences in the continuous transition showed on Fig. 2. Their sequence revealed that the pattern 1, 2, 3, 8… is a transient condition between two successive expressions of the main Fibonacci pattern: 1 : 2 and 2 : 3 (Fig. 6). The divergence value 144, typical for the 1 : 2 expression (in orthogonal lattice) drops down slightly below 130 before rising again towards 138.46 for 2 : 3 and approaching finally 137.5 for higher expressions. In this area the primordia are tightly packed with very little space left between them. This results in the most efficient use of the organogenic surface. It became immediately clear that if the primordia acquired DM in this particular region, the pattern would become ‘petrified’ – it would transform no more in the simulation, but would be perpetuated as in the case shown in Fig. 5 on the left. When DM is reached in the higher up region – the main Fibonacci pattern is a result. The PA initial condition determines then where the ultimate value of R is going to be reached.
|
It remains to be seen whether the two options predicted by the model occur in nature. The good plant system for investigating it is an ancient woody magnoliid – magnolia. Its vegetative phyllotaxis is known to transform readily from initial distichous to helical one. This research is currently under way.
In the unlimited continuous transitions with 1 : 1 IP, the 1, 2, 3, 8, 11… pattern rises from and transforms into the main Fibonacci pattern (Fig. 6). For now, it is unclear how to obtain, if possible at all, a progression from its low 2 : 3 expression towards the higher ones: 3 : 8 or 8 : 11, encountered in magnolia or balsam fir. These have also been found in Annanas comosus – a monocotyledonous plant with showy multiple fruit (Rutishauser and Peisl 2001; B. Zagórska-Marek, unpubl. data). In all these plants, the high expressions of the pattern discussed resulted from transitions involving patterns different that the main Fibonacci – tetrajugy (4 : 8) or Lucas (7 : 11). Further investigations are needed both of the nature of this pattern and of the conditions of its appearance in transitions.
It is evident from our study that the quality of phyllotactic pattern resulting from transition depends not only upon the relation of primordia sizes relative to the size of organogenic surface, but also upon how the emerging phyllotactic lattice is rotated with respect to the axis of the system, and how efficiently the primordia are packed.
The parastichy orientation in a continuous transition puts in question the identity of phyllotactic pattern. In general, we believe that the problem of phyllotactic transitions will bring us soon to redefinition of some patterns and perhaps to their new classification.
Discontinuous change
Alternatively, the change of R in our simulations was abrupt. This is what usually happens when SAM enters a new phase of development and the primordia assume new identity: instead of being leaf primordia they become the primordia of inflorescence or flower elements. We have used two model cases to illustrate this type of transition and confront the simulation results with experimental data. The first case refers to the transition from vegetative to generative phase of development (case study Verbena), and the second to the multiple transitions between the primordia of floral parts (case study Magnolia).
Case study: Verbena
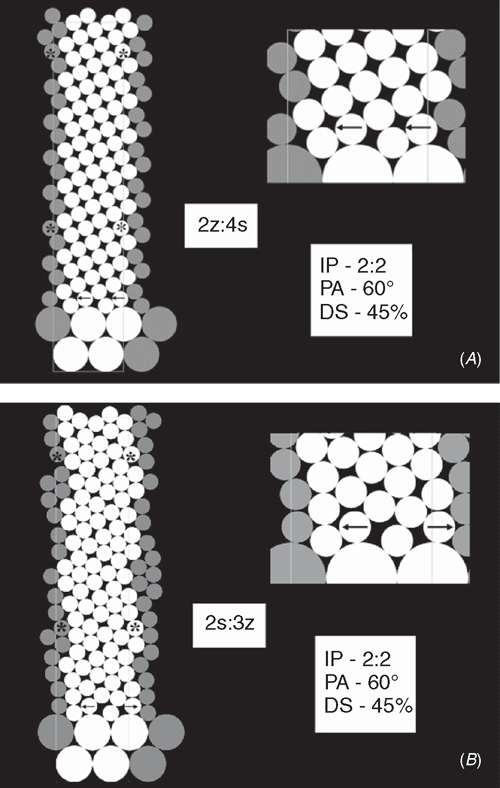
In Verbena officinalis, vegetative phyllotaxis is decussate (achiral), whereas on the inflorescence axis the bract distribution is helical (chiral). The transition from the decussate arrangement to the helical one, most commonly leads to the development of the main Fibonacci pattern. The frequent alternative is bijugy. The youngest (uppermost) primordia of leaves are larger, relative to the SAM circumference, than the primordia of bracts. Modelling this situation showed quickly the ambiguity of the first available space rule when the difference between the sizes of the primordia was sufficiently large. The DS parameter, a discontinuous percentage change in R, had to be smaller than 50%. In the simulations with DS = 45%, after the change, all new primordia were small and all were the same size. The quality of the emergent pattern depended upon how the small primordia were positioned in quite a large available space. When, in a case of two equivalent positions, the two small, ‘generative’ primordia had their initiation site selected identically – to the right or to the left – the developing pattern was bijugy, when the choice for both had been opposite – one to the left, the second to the right – the main Fibonacci pattern emerged (Fig. 7). The latter transition should be the most frequent among plants, quickly developing helical phyllotaxis from the initial pattern of opposite cotyledons or prophylls.
|
In a sense, this case resembles the one shown in Figs 1 and 2 except that there, with an initial patten 1 : 1, ambiguity of the first available space rule resulted in two chiral configurations of the same pattern, but here, the number of possible patterns is greater (including their chiral variants). This increases phenotypic plasticity of the plant: it is definitely better to have two cotyledons than one.
Case study: Magnolia
The outstanding diversity of phyllotactic patterns present in the terminal parts of magnolia flowers (Zagórska-Marek 1994; Zagórska-Marek and Wiss 2003) is most probably an outcome of the double change in the size of floral primordia generated on SAM’s organogenic lateral surface (Zagórska-Marek and Stoma 2005). The computer simulation of this case is very sensitive even to small changes in the values of the parameters used. The ambiguity of the first available space rule is also of importance here. The initial pattern of the largest perianth primordia is tricussate (achiral), which explains frequent appearance of trijugy in the later helical arrangement of the smaller stamen primordia and slightly larger carpel primordia (Fig. 8). In the simulation, very small primordia of stamens, again, have a choice of more than one position in the first available space, identically like in the case of Verbena. There are two variants of this situation: (1) asymmetric, when two small primordia are getting close one to another with the third one being far apart from the pair, or (2) symmetric, when all three are shifted in the same direction thus are equally spaced. This might be one of the reasons why helical phyllotaxis of generative elements in magnolia is extremely diverse, covering almost whole spectrum of patterns known to occur in nature. Frequent ontogenetic transitions through dislocations (bifurcations of parastichies) are observed in a carpel zone (Zagórska-Marek 1994). A tricussate start up pattern, together with rather small size of generative elements in relation to the width of organogenic space (ambiguity of available space rule), but above all, the double change in a size of pattern elements should be held responsible for the rich spectrum of final phyllotactic solutions. In our simulations, we have sometimes observed the development of totally chaotic arrangements of pattern elements. Screening large populations of magnolia generative shoots shows that this situation is also present in nature (Zagórska-Marek 1994). Frequent appearance of parastichies with a somewhat irregular course in silico and in vivo (Figs 8 and 9) explains why, in some cases, phyllotaxis of magnolia gynoecium is undeterminable, with only one set of parastichies clearly helical (B. Zagórska-Marek, unpubl. data). Similar effects have been observed in silico in tiling patterns of phyllotaxis by Atela and Gole (2007) and earlier, both in silico and in vivo, by Couder (1998) for Helianthus giving, according to the latter author’s opinion, some substance to the Plantefol’s claim on existence of ‘foliar helices’ (Plantefol 1948).
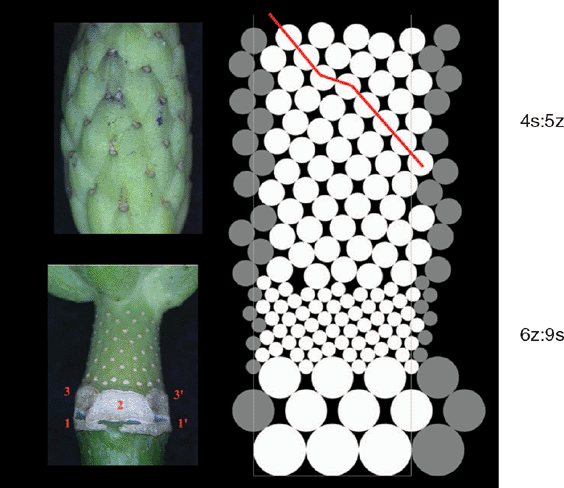
|

|
Conclusion and future goals
Our simulation results coincide well with the real organographic changes in the systems of phyllotaxis exemplified by Magnolia and Verbena shoots. Other authors observed similar concordance using different plant model systems (Couder 1998; Hotton et al. 2006; Smith et al. 2006b; Hellwig et al. 2008). The new development in our model application is that, sometimes, there is more than one initiation site available for an emerging primordium. Its selection, together with other factors such as the varying extent and rate of change in primordia size, affects the quality of the resulting pattern and ontogenetic transition. We note that, in some plants such as Dipsacus or Torreya, the pattern subsequent to decussate is always bijugy, suggesting that there is yet another factor, besides the primordia’s competition for auxin, which selects arbitrarily the initiation site for new primordium. This observation may put in question the absolute autonomy of SAM in its organogenic activity (Banasiak and Zagórska-Marek 2006).
Still, some unexplained problems remain. Firstly, qualitative changes in phyllotaxis sometimes occur within the zone of primordia of the same identity. In Verbena, on the axis of inflorescence, further sequential changes of phyllotaxis often take place even though the identity of bract primordia remains the same. The transitions we have modelled (data not shown here since this research is still in progress) involve helical patterns (Lucas, second accessory) and whorled (tricussate); similarly, in Magnolia flowers, dislocations, frequently observed in a carpel zone, rearrange the primordia, which maintain their identity. Of the same type are the transitions in A. balsamea vegetative shoots, that occur within one annual increment of the axis (Zagórska-Marek 1985). Secondly, the spectrum of final patterns in Magnolia is species or genet specific, despite the general rule of a double change in primordia size of flower elements. Different Magnolia species, or even the trees within one species, may have specific distribution of various phyllotactic patterns. This ‘phyllotactic fingerprint’ can be maintained throughout the period of many years (Zagórska-Marek 1994, 1999). Truly challenging are the species having exclusively main Fibonacci pattern in the carpel arrangement (e.g. Magnolia hypoleuca, Magnolia virginiana). Evidently the unknown species or genet specific factor controls the direction of transitions in magnolia’s floral phyllotaxis.
Our future specific goals in exploring the potentials of the presented model in more detailed, quantitative studies, are to explain systematically and predict all (including the less common) patterns, explain the direction of other pattern transitions, i.e. why a given pattern may be transformed in many different ways, and, finally, elucidate why the spectrum of patterns can be species or genet specific.
Acknowledgements
B. Zagórska-Marek thanks her colleagues from the Plant Development Research Group at the Institute of Plant Biology of Wrocław University for their interest in phyllotaxis and great help in counting and gaining control over thousands of plant spirals, especially to Ms Magdalena Turzańska for beautiful microtechnics performed on shoot apical meristems. Thanks are also due to computer programmers: Dr Radosław Karwowski and Szymon Stoma for their efforts to improve the first, unfinished version of the computer program, meant to allow simulations of phyllotactic transitions. It was written in 1995 by Dr Johannes Battjes during our cooperation in Professor Prusinkiewicz’s Laboratory at the University of Calgary. This research was financed in part by a grant from the Polish Ministry of Science and Higher Education (Grant no. N303 096834).
Adler I
(1974) A model of contact pressure in phyllotaxis. Journal of Theoretical Biology 45, 1–79.
| Crossref | GoogleScholarGoogle Scholar | PubMed |

Adler I
(1977) The consequences of contact pressure in phyllotaxis. Journal of Theoretical Biology 65, 29–77.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Banasiak A, Zagórska-Marek B
(2006) Signals flowing from mature tissues to SAM determine the phyllotactic continuity in successive annual increments of the conifer shoot. Acta Societatis Botanicorum Poloniae 75, 113–121.
Couder Y
(1998) Initial transitions, order and disorder in phyllotactic patterns: the ontogeny of Helianthus annuus. A case study. Acta Societatis Botanicorum Poloniae 67, 129–150.
Fujita T
(1937) Ueber die Reihe 2,5,7,12,… in der schraubigen Blattstelung und die matematische Betrachtung verschiedener Zahlenreihensysteme. Botanisches Magazine Tokyo 51, 480–489.
Fujita T
(1939) Statistische Untersuchungen ueber den Divergenzwinkel bei den schraubigen Organstellungen im Pflanzenreich. Japanese Journal of Botany 12, 1–55.
Heisler M,
Ohno C,
Das P,
Sieber P,
Reddy G,
Long J, Meyerowitz E
(2005) Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Current Biology 15, 1899–1911.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hellwig H,
Jeschke M, Deussen O
(2008) The dynamic generation of compact phyllotactic patterns on surfaces of revolution. Journal of Theoretical Biology in press. ,
Hotton S,
Johnson V,
Wilbarger J,
Zwieniecki K,
Atela P,
Gole C, Dumais J
(2006) The possible and the actual in phyllotaxis: bridging the gap between empirical observations and iterative models. Journal of Plant Growth Regulation 25, 313–323.
| Crossref | GoogleScholarGoogle Scholar |
Jönsson H,
Heisler M,
Shapiro B,
Meyerowitz E, Mjölsness E
(2006) An auxin-driven polarized transport model for phyllotaxis. Proceedings of the National Academy of Sciences of the United States of America 103, 1633–1638.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Larson P
(1977) Phyllotactic transitions in the vascular system of Populus deltoides Bartr. as determined by 14C labelling. Planta 134, 241–249.
| Crossref | GoogleScholarGoogle Scholar |
Liaw S
(1998) Phyllotaxis: its geometry and dynamics. Physical Review E: Statistical Physics, Plasmas, Fluids, and Related Interdisciplinary Topics 57, 4589–4593.
Reinhardt D,
Mandel T, Kuhlemeier C
(2000) Auxin regulates the initiation and radial position of plant lateral organs. The Plant Cell 12, 507–518.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Reinhardt D,
Pesece E,
Stieger P,
Mandel T,
Baltensperger K,
Bennett M,
Traas J,
Friml J, Kuhlemeier C
(2003) Regulation of phyllotaxis by polar auxin transport. Nature 426, 255–260.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Smith RS,
Guyomarch S,
Mandel T,
Reinhardt D,
Kuhlemeier C, Prusinkiewicz P
(2006a) A plausible model of phyllotaxis. Proceedings of the National Academy of Sciences of the United States of America 103, 1301–1306.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Smith RS,
Kuhlemeier C, Prusinkiewicz P
(2006b) Inhibition fields for phyllotactic pattern formation: a simulation study. Canadian Journal of Botany 84, 1635–1649.
| Crossref | GoogleScholarGoogle Scholar |
Snow M, Snow R
(1931) Experiments on phyllotaxis. I. The effect of isolating a primordium. Philosophical Transactions of the Royal Society of London 221B, 1–43.
Snow M, Snow R
(1952) Minimum areas and leaf determination. Proceedings of the Royal Society 139B, 545–566.
Williams RF, Brittain EG
(1984) A geometrical model of phyllotaxis. Australian Journal of Botany 32, 43–72.
| Crossref | GoogleScholarGoogle Scholar |
Zagórska-Marek B
(1985) Phyllotactic patterns and transitions in Abies balsamea. Canadian Journal of Botany 63, 1844–1854.
Zagórska-Marek B
(1987) Phyllotaxis triangular unit; phyllotactic transitions as the consequences of the apical wedge disclinations in a crystal-like pattern of the units. Acta Societatis Botanicorum Poloniae 56, 229–255.
Zagórska-Marek B
(1994) Phyllotaxic diversity in Magnolia flowers. Acta Societatis Botanicorum Poloniae 63, 117–137.